 Mixing
Mixing
カメ
この記事の正確性に疑問が呈されています。問題箇所に信頼できる情報源を示して、記事の改善にご協力ください。議論はノートを参照してください。(2012年8月) |
| カメ目 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ドイツの生物学者エルンスト・ヘッケルが著した『自然の芸術的形態(Kunstformen der Natur)』(1904年刊)の中の1図 数字は画像ページのものに対応 1.オサガメ 2.タイマイ 3.ギザミネヘビクビガメ 4.マタマタ 5.ホシヤブガメ 6.アルダブラゾウガメ 7. カミツキガメ | |||||||||||||||||||||
地質時代 | |||||||||||||||||||||
| 約2億1,000万年前(中生代三畳紀後期) - 新生代第四紀完新世(現世) | |||||||||||||||||||||
分類 | |||||||||||||||||||||
| |||||||||||||||||||||
学名 | |||||||||||||||||||||
Testudines Linnaeus, 1758 | |||||||||||||||||||||
和名 | |||||||||||||||||||||
| カメ目 | |||||||||||||||||||||
| 英名 | |||||||||||||||||||||
| Testudines | |||||||||||||||||||||
亜目,属 | |||||||||||||||||||||
|
カメ(亀、ラテン語名:Testudo、英語名:Turtle, etc.)[要検証]は、爬虫綱-(双弓類[1])- カメ目(亀目、学名:Testudines、英語名:Testudines)に分類される爬虫類の総称。絶滅した初期グループおよび、現存する曲頸亜目・潜頸亜目の2亜目で構成される。
現生と化石の別無く、すべての種が胴体を甲羅としている点が特徴となっている。しかし、2018年8月、中国貴州省にて、甲羅のないカメのほぼ完全な化石が発見されたとネイチャーにて発表された。これにより、古代のカメには甲羅がなかったことが明らかになった[2]。
目次
1 概説
2 形態
2.1 大きさ
2.2 二次性徴
2.3 頸
2.4 頭部
2.5 甲羅
2.6 皮膚と脱皮
2.7 四肢
2.8 卵
3 生態
3.1 長寿記録
4 分類
4.1 上位分類
4.2 下位分類
5 進化
5.1 三畳紀のカメ
5.2 曲頸類と潜頸類の出現
5.3 ウミガメの出現
6 分布
6.1 陸地
6.2 海洋
6.3 日本で見られる種
7 利用
7.1 食用
7.2 甲羅の利用
8 ペットの亀
9 文物に登場する亀
9.1 神話・伝説の亀
9.2 吉兆・縁起物としての亀
9.3 「動きの遅いもの」としての亀
9.4 侮蔑表現としての亀
9.5 近現代の創作の題材としての亀
10 呼称
10.1 学名(ラテン語)
10.2 漢字文化圏
10.3 英語
10.4 ドイツ語
10.5 タイ語
11 亀に関する言葉
11.1 カメの名を持つ生物
11.1.1 和名
11.1.2 英名
12 画像
13 脚注
14 参考文献
15 関連項目
16 外部リンク
概説
カメ目は、多様な爬虫類グループの中でも比較的早期の約2億1000万年前(中生代三畳紀後期)に出現し、甲羅を本格的に発達させたことで特徴づけられる一群で、現代まで継続して繁栄している。ヒトの出現、乱獲によって絶滅した種、あるいは危惧される種があるが、それでも、グループ全体としては水・陸の両域で多様性を維持している。
文化的側面で言えば、亀は、堅い守りの象徴である一方で鈍重を揶揄する語として使われる。また、歩みの遅さではなく着実さを肯定的に捉える場合には、実直さや勤勉さなどを褒めるための比喩となる。古来、中国やインドでは神獣として扱われ、中国文化の影響下にあった地域では吉兆とされる。
形態

甲羅と四肢の繋がり
基本的な構造は、四肢動物の基本から大きく逸脱するものではない。ただし、その胴部がはっきりした甲羅を構成する点が最大の特徴となっている。これは、内部構造では脊椎骨、肩胛骨、肋骨、胸骨などが互いに密着して箱のような構造をなしている。また外側ではブロック状に並んだ板によって外見的な甲羅が形成されるが、これは二次的に退化したものもある。甲羅は腹面、背面、側面で閉鎖されており、前側の窓から頭部と前足、後ろ側の窓から後ろ足と尾が出る形になっている。このように四肢帯が肋骨に囲まれているのは脊椎動物ではこの類以外になく、現生爬虫類中でもっとも特殊化した形態である[3]。
大きさ

亀の大きさはこのように背甲の直線距離で測る(甲長)
本目の構成種は甲羅に頸部や尾を収納する種が多いため、頭胴長(体長)や全長を測ることが難しい[4]。そのため背面の甲羅(背甲)の直線距離(背甲長、単に甲長とも)で大きさを表す[4]。現生の最大種はオサガメで最大甲長183センチメートル以上[5]。最小種はシモフリヒラセリクガメで最大甲長9.6センチメートル[6]。
大型種としては、コガシラスッポン属(インドコガシラスッポンは甲長140cm)、アルダブラゾウガメ(甲長120cmで体重300kg)、現生最大のウミガメであるオサガメ(甲長約200cm、重さ900kg以上)などを挙げることができる。
過去の絶滅種には全長4mに及ぶウミガメであるアーケロン(Archelon spp.。最大甲長1.9m。中生代白亜紀、米国)や、最大甲長でそれを上回る淡水棲のヨコクビガメ類であるスチュペンデミス・ゲオグラフィクス(Stupendemys geographicus。全長約4m、最大甲長約2.35m、最小甲長約1.8m。新生代中新世トートニアン、ベネズエラ)などの大型種が存在した。有史以前にはリクガメ属[要検証]の仲間やメイオラニアなど、2.5mを超える種が世界中の比較的広い範囲に分布しており、南北アメリカやオーストラリア、アフリカなどに棲息していたことが知られている。
二次性徴
オスとメスの大きさは同じか、多くの種ではメスのほうが大型化する[要検証]。極端な例としてはカンムリガメが挙げられ、17.5cmにとどまるオスの最大甲長に対し、メスでは61cmに達する。主な二次性徴として、オスは陰茎を収納する関係から尾が太く長く、交尾の際メスに乗りやすいよう腹甲が凹んでいる種が多い。
頸
多くの種は頸椎を垂直または水平方向に曲げることで頭部を甲羅に収納できるが、頭部を甲羅に収納することができない種もいる[7]。
原始的カメであるプロガノケリス類をはじめとして最初期に棲息していたカメ類は、頭部を収納できず、棘で武装するという違う方向性での進化を見せていた。しかし、その後のカメ類の進化は頭部と四肢を甲羅に中に納める方向に向かった。
ここで、収納を実現するために曲頸類のカメがとった方法は、上下の甲羅の間に頸を横向きにして折り畳み、挟み込むように納める「曲頸(きょくけい)」である。しかし、曲頸では、収納に成功してはいても頭部と頸部の半分が露出しており、完全に保護できているわけではない。
一方、遅れて出現した潜頸類のカメがとった方法は、頸を垂直方向にS字形に湾曲させて文字どおり中に押し込めるように収納する「潜頸(せんけい)」である。世界中の棲息地で曲頸類は潜頸類に取って代わられていった。
海棲に進化したものは、海に適応するにしたがって頭部を収納する必要がなくなり、退化的進化を遂げたものである。進化については、「#進化」も参照。
オオアタマガメは潜頸類であるが、巨大な頭部を持つため、それを甲羅に納められない。また絶滅種であるメイオラニアも頭部を甲羅に納められなかった。

リクガメの頭部

ヒガシトゲスッポンの頭部

| 背甲 (左図) | 腹甲 (右図) |
| 項甲板 | 1 間喉甲板 |
| 椎甲板 | 2 喉甲板 |
| 肋甲板 | 3 肩甲板 |
| 縁甲板 | 4 胸甲板 |
| 臀甲板 | 5 腹甲板 |
| 6 股甲板 | |
| 7 肛甲板 | |
| 8 腋下甲板 | |
| 9 下縁甲板 (2x) | |
| 10 鼠蹊甲板 |

甲羅にはいった子ガメ(カロリナハコガメ)

リクガメ(イベラギリシャ)の甲羅

水棲ガメ(クサガメ)の甲羅
頭部
水棲カメの眼は横側についているものも多いが、陸上で暮らすカメの多くは、対象を見下ろせるように眼が前方についている[要検証]。カミツキガメやスッポンのような水棲カメの一部では、頭の上部付近についている。これらの種は、浅い水の中に眼と鼻孔を除く体全体を潜めて天敵から身を隠すことができる。ウミガメは、眼の近くに塩分を含む涙を作る涙腺を具えていて、飲んだ海水から得た体内の過剰の塩分を排出することができる。カメの網膜には、通常ほかの生物に見られるよりも多くの桿体細胞があるため、非常に夜目が利くと考えられている。また、近紫外線(UV A)から赤にわたる範囲に感度がある錐体を持っている。
草食性のリクガメの一部は、素早く動く餌を追跡して狩るための俊敏さが無いが、肉食性のカメは、頭部を素早く振ることができる。現生種では歯がなく、顎は角質の鞘(嘴)で覆われる[5][8]。この硬い嘴(くちばし)を具えた顎を用いて餌を切断したり咀嚼したりする。餌を呑み込むためにカメは舌を用いるが、トカゲやヘビなどの他の多くの爬虫類に見られるように舌を突き出してペロリと食物を捉えることなどはできない。植物食の種では硬い植物を噛みきれるように嘴に鋸状の突起がある種や[5]、動物食の種では獲物を切断できるように嘴が薄く刃物状になった種もいる[8]。また嘴が幅広く、硬い食物を噛み砕くことができる分類群もいる[5][8]。
甲羅
カメの形態上の最大の特徴は、甲羅を持つことである[8]。甲羅は脊椎や肋骨と一体の甲板(骨甲板)と、鱗からなる甲板(角質甲板)の2つの甲板で構成される[5][8][9]。腹甲の一部は鎖骨や肋骨が変形したとされる[5]。骨甲板と角質甲板の継ぎ目がずれており、強度をあげている。
カメの甲羅(骨甲板)は肋骨や背骨のみが変形してできたとする説と、肋骨や背骨が「皮骨」と融合してできたという説とがあった。しかし2013年、理化学研究所は、カメの胚の発生プロセスの組織学的な解析と三畳紀の化石の調査により、カメの骨甲板は肋骨だけが拡張・変形して進化してきたことを立証したと発表した。ワニやアルマジロなど他の脊椎動物の装甲は、真皮層で形成された皮骨という組織からなるが、カメの背甲の板状の骨は肋骨が作られたのち骨膜が拡張し、その骨膜内で形作られるもので、その形成は真皮より下の結合組織内で起きる[10]というものである。
角質甲板は以下のように多くのパーツから構成されている[5][8]。分類群によってこれらの有無や数が決まっているが、発生時の環境や外傷、疾病などにより奇形を生じることもある[8]。
- 背甲(carapace)
- 背面にある甲羅。
- 項甲板(nuchal、precentral、または、cervical)
- 背甲の頭部側の先端にある左右の縁甲板をつなぐ甲板。これが無いカメもいるので、識別の重要なポイントになり得る。
- 椎甲板(vertebral、または、central)
脊椎の上部にある甲板。多くのカメでは5枚。前から第一椎甲板、第二椎甲板、 ...書 き か け... 、第五椎甲板と呼ぶ。- 肋甲板(pleural、costal、または、lateral)
- 背甲の肋骨(人間と違い、肩帯から腰帯まで覆う)上部にある甲板。椎甲板の左右に4対ある。
- 縁甲板(marginal)
- 背甲(背面だけでなく腹面も含めて)の外縁を覆う12対ある甲板[要検証]。ワニガメでは肋甲板との間に上縁甲板があり原始的な特徴とされる。分類群によっては最も後部にある臀甲板が癒合し1枚(臀甲板と呼称されることもあるが一般的でない)になる。
- 臀甲板(prostcentral、caudal、または、supracaudal)[要検証]
- 背甲の最も尾側にある縁甲板。第十二縁甲板は特別に臀甲板と呼ぶ。種類によって、左右1対か融合して1枚になっている。臀甲板が1つのことを「第十二縁甲板は融合している」、2つのことを「第十二縁甲板は分かれている」ということがあり、種の識別に役立つ。

ギリシャリクガメの第十二縁甲板は融合している

クサガメの第十二縁甲板は分かれている
- 上縁甲板(supramarginal)[要検証]
- ワニガメには肋甲板と縁甲板の間に上縁甲板(supramarginal)がある。
- 上縁甲板(supramarginal)[要検証]
- 腹甲(plastron)
- 腹面にある甲羅。いくつかの属や種では甲板の間に1、2つの蝶番があり可動させることができる。
- 間喉甲板(intergular)
- 喉甲板の上か間にあり、肩甲板、胸甲板の間にある場合があり、ヨコクビガメ科、ヘビクビガメ科などに ----( 書 き か け )---- 。
この節の加筆が望まれています。 |
- 喉甲板(gular)
- 腹甲のうち、一番頭部に近い位置にある左右に1対の甲板。左右の喉甲板が癒合し1枚になった分類群もいる。種類により1枚の場合と2枚の場合がある。
- 肩甲板(humeral)
- 前足の付け根に近い位置にある左右に1対の甲板。
- 胸甲板(pectral)
- 前足の後ろの位置(胸)にある左右に1対の甲板。
- 腹甲板(abdominal)
- 股甲板(femoral)
- 肛甲板(anal)
- 腹甲のうち、一番尾に近い位置にある左右に1対の甲板。種によってはこの甲板の間にある切れ込みにより、雌雄を判別できる。
- 橋(きょう、bridge)
- 背甲と腹甲の間の部位。上記の胸甲板や腹甲板が外側へ張りだし、縁甲板と接している分類群が多い。
- 下縁甲板(inframarginal)[要検証]
- 縁甲板と胸甲板の間、橋の前肢の基部後方にある甲板
- 腋下甲板(axillary)
- 腋(わき)の下、前足の付け根の甲板。
- 鼠蹊甲板(ingunal)
- 縁甲板と腹甲板の間、橋の後肢の基部前方にある甲板。鼠蹊部、すなわち、後ろ足の付け根の甲板。
- 下縁甲板(inframarginal)[要検証]
現生種では化石種と比較して甲板が薄く軽量化し[5]、甲板数も少ない傾向がある[8]。例外もあるが陸棲傾向の強い種では甲板が分厚く背甲がドーム状に盛り上がり、水棲傾向の強い種では水の抵抗を減らすため甲板が薄く背甲が扁平になる傾向がある[5][8]。一方で陸棲種ではパンケーキガメのように非常に甲羅が扁平で、素早く隙間にもぐりこむ種もいる[5][8]。水棲種ではフロリダアカハラガメやマレーハコガメの亜種などのようにドーム状に盛りあがる背甲を持つ種もいて、これは同所的に分布するワニなどの捕食者に対する防衛手段(甲羅が分厚くなることで飲みこみにくくなる)と考えられている[5][8]。陸棲種では腹甲が大型(背甲よりも長いことが多い)になり、水棲種では腹甲が小型になる傾向がある[8]。スッポン上科やオサガメは軽量化のため角質甲板が無く骨甲板も退化しているが[5][8]、これは浮力により体重を支えることができ表面を甲板ではなく皮膚で被うことで水の抵抗を減らす効果があると考えられている[9]。例外も存在し、陸棲でありながら扁平な背甲を具え素早く動き岩の隙間等に潜り込むパンケーキガメや、水棲傾向が強いがドーム状の背甲を持つマレーハコガメの亜種などがいる。複数の科において腹甲に蝶番状の機構がある分類群が存在し、これにより腹甲を折り曲げて可動することができる[5][8]。蝶番のある多くの分類群で1か所、ドロガメ属のみ2か所、セオレガメ属のみ背甲に蝶番がある[8]。蝶番による腹甲の可動する利点としては背甲と腹甲の隙間を減らすことによる外敵や乾燥からの防御、逆に背甲と腹甲の隙間を増やすことで大型の卵を産むことができる(幼体や栄養分の増加により死亡率を減らせる)などの効果があると考えられている[5][8]。
なお、甲羅を持つがゆえに他の生物には見られない特徴が見られる。本来、肩帯は、肋骨が存在している場合は胸郭の肋骨より外側に付くのが普通である。しかし、カメでは発生時に肋骨が外側に広がり肩帯を取り込む[7]ため、四足動物では本目のみ肋骨(甲羅)の内側に肩帯がある[5][4][7]。また、肘関節は他の爬虫類とは逆に外側に曲がるようにできている。腰帯は他の四肢動物と同じく仙椎を介して体幹に繋がっているが、仙椎直前の胴胸椎肋骨と癒合した肋骨板が仙椎と癒合した上尾骨板とともに後方に伸長して甲羅後縁を形成するため、結果として甲羅の内側に位置する。また、かつて胴体を動かしていたと思われる筋肉は、甲羅により胴体を曲げたりしなくなったため退化したように見えるが、肺呼吸をするうえで鞴(ふいご)のように働き、呼吸運動に必要な力を供給する隔膜として転用されている。ただし、隔膜は人間の横隔膜と違い、縦に付いている。カメは主に肺呼吸を行うが、肺は大型であるものの胴体が甲羅で覆われているため、胸筋や腹筋を使って肺を収縮・膨張させて呼吸することはできない[8]。そのため頭部や四肢を甲羅に入れることで肺を収縮させ、肺の中の空気を吐き出し、逆に頭部や四肢を甲羅から出すことで肺をふくらませ空気を吸いこむ[5][8]。水棲種では鼻や口、喉の粘膜、総排泄口にある粘液嚢、皮膚を使い、副次的ではあるが皮膚呼吸によるガス交換をおこなう種もいる[8]。
また、カメの甲羅は、カリウムやマグネシウムといった栄養素を貯蔵する役割も持つ[2]。
皮膚と脱皮
前述のように、甲羅の外側の層は皮膚の一部からできていて、甲板は一枚一枚が鱗に相当する。甲羅以外の皮膚の残りの部分は、他の爬虫類のようなそれよりも小さい鱗から成る皮膚で構成される。
陸棲種では皮膚が大型の鱗で覆われ[4]、乾燥から身を守っている。水棲種では皮膚に大型の鱗がある種は少ない(陸上でも水中にも生息する種はこのかぎりではない)が、これにより皮膚呼吸が可能になり水中での活動時間を長くしている。
水棲カメはヘビのように全部を一度に脱皮することはせず、小さい部分ごとに行う。水中にいるときは、カメが木や石などに体をこすりつけて、皮の薄いシート(薄い樹脂フィルムのかけらのように見える)を剥がす様子が観察される。リクガメも脱皮を行うが、死んだ皮膚は山状に積層することがあり、それが甲羅の外側の部分を保護するのに役立つ。

歩行するリクガメ

水陸両棲のカメの四肢

アオウミガメの鰭状の四肢
一年でおよそどれだけの鱗甲ができるのかを知っていればカメの甲羅の積層によってできるリング模様を数えることで、およそのカメの年齢を見積もることができる[11]。ただし、カメの成長速度は一定ではない上に、鱗甲の一部はときどき剥落するので、この方法はあまり正確ではない。
四肢
リクガメは、短くて丈夫な足を持っている。リクガメは古来より歩みが遅いものとして認識されているが(#「動きの遅いもの」としての亀参照)、それは重くて邪魔になる甲羅があるため(甲羅を軽量化し素早く隙間にもぐりこむパンケーキガメといった例外もある)と、足が哺乳類のように体の下側にまっすぐ伸びているわけではなく、トカゲのように足が曲がってついているため、歩行運動の効率があまりよくないからである。
陸棲種では指趾が退化し[5]、鉤爪が発達する。水棲種は逆に指趾が長く、その間に水かきが発達する[5][8]。スッポンモドキやウミガメ上科では指趾の境目が不明瞭で、四肢はひれやオール状になる[5]。
ウミガメは前肢を上下運動させて推進力を得て、あたかも水中をはばたいて飛ぶように泳ぐ。後肢は推進には使われず、舵の役割を果たしている[5]。孵化(ふか)したての幼カメが巣から海まで移動するとき以外の場合は、淡水棲カメと比較するとウミガメは地上での動作がとても制限されている。メスのウミガメは産卵のために再上陸しなければならないが、そのときはひれ状の四肢を用いて苦労して体を前に引きずって前進する。
水陸両棲のカメは通常、リクガメによく似た四肢を具えているが、多くの種がアヒルのように指趾の間に水かきをもつ。水陸両棲のカメは四肢をすべて使って犬掻きに近い方法で泳ぐ。右側の2本の足と左側の2本の足を交互に前後させる。ワニガメのように、湖沼や河川の底をただ歩くことを好む種も存在する。ヌマガメ科の一部では、オスのカメは、メスに比べて特に長い鉤爪を持っている傾向がある。これは、交尾をする際にメスをつかまえておくための特徴であると考えられている。スッポンモドキのように、鉤爪が比較的小さくなり、指と指の間がつながった完全な水かきを持つカメもいる。これらのカメは前述のウミガメと同じ泳ぎ方をする。
卵
卵はウミガメ科、カミツキガメ科、スッポン科では球形、ドロガメ科やヌマガメ科では楕円形で、他科では種によって異なる[5]。主にウミガメ科、カミツキガメ科、ヨコクビガメ科は皮革状の柔らかい殻、主にドロガメ科、スッポン科、ヘビクビガメ科では鶏卵状の硬い殻で被われ、他科では種によって異なる[5]。
生態
淡水域、海洋、砂漠、草原、森林などの様々な環境に生息する[7]。緯度の高い地域に生息する種では氷の張った水面下での活動が観察されている種もいる。樹上棲種はいないが、オオアタマガメは四肢や尾を使い多少ながら木に登ることもある[7]。陸棲種のみで構成されるリクガメ科から、産卵を除いて上陸しない完全水生種(海棲種)のみで構成されるウミガメ上科もいる[8]。多くの種類が河川、湖沼、池等の淡水域に生息する[要検証]。水からあまり離れずに生活するが、リクガメ科は終生陸上で生活する。水棲種も多くは肺呼吸をしないと生きていけず、水面に顔を出して息継ぎを行う。しかし、冬眠中の個体やハヤセガメのように総排出腔でガス交換を行うことにより空気呼吸を行わず、呼吸のために水面に上がらない種もいる。
食性は種によって異なり[7]、魚類、両生類、爬虫類、哺乳類、昆虫、貝類、甲殻類、植物の葉、果実、キノコなどを食べる種が知られている。一部の食物を専食する種もいる。水棲種では口を開け舌弓を動かし喉を広げることで口内の水圧を低下させ、水ごと獲物を吸い込み捕える事が多い[5][8]。
繁殖形態は卵生。主に陸上に産卵するが[5]、浅い水中に産卵(発生は水没していない状態で進む)する種もいる[8][12]。海を広範囲にわたって回遊するウミガメ類も産卵時は砂浜に上陸する。エミスムツアシガメは産卵巣の上に木の枝や落ち葉を塚状に積み上げ、母親が卵を保護する[5]。
長寿記録
カメ類は細胞の代謝のサイクルが遅く[要検証]、動物の中でも長寿の代表格とされる。
確実な長寿記録として1766年にセーシェルからモーリシャスに持ち込まれ、1918年に死亡したアルダブラゾウガメ(マリオンのゾウガメ)の152年の飼育記録がある[5][9]。他種ではギリシャリクガメ(ティモシー)の1855-2004年にかけて149年(1842年生まれの162歳だったとされるが根拠は不明)、カロリナハコガメの138年、ヨーロッパヌマガメの120年の記録がある[9]。
不確実な記録としては1835年にダーウィンに採集され2006年に死亡したサンタクルスゾウガメ(ハリエット)は175年の飼育記録があるが、ハリエットはダーウィンが上陸しなかった島に分布するサンタクルスゾウガメであることが判明したためダーウィンが採集した個体ではないとする説もある[9]。1750年生まれとされアリポーア動物園で2006年に死亡したアルダブラゾウガメ(アドワイチャ)の255年の記録があるが、飼育していたとされるロバート・クライブが最後にインドにいた1767年からアリポーア動物園が開園する1875年までの記録がないこと、1875年にセーシェルから持ち込まれた個体とする報道もあり不確実とされる[9]。1773年か1777年にジェームズ・クックがトンガの女王に送り1966年に死亡したホウシャガメ(トゥイ・マリリア)の189-193年の記録があるが、ジェームズ・クックがホウシャガメの分布するマダガスカルに寄港歴がないこと、ジェームズ・クックおよびトンガの双方にもホウシャガメの譲渡に関して記録がないことから不確実な記録とされる[5][9]。
分類
上位分類
カメ目の分類には、紆余曲折の歴史がある。従来、四肢動物の大まかな分類は、側頭窓(temporal fenestra)の形態によって行われていた[13]。20世紀初頭、オズボーンによって提案された当時の分類では、側頭窓を一つだけもつかあるいは持たないものが単弓類とされ、カメはここに分類された。その後、無弓類、広弓類などが提案され、カメはパレイアサウルス類などとともに無弓亜綱とされた[8][7][14]。しかし近年は形態や発生学の研究から双弓亜綱に含まれるとする説が有力で[8]、さらに分子系統学的解析から主竜類もしくは主竜形類に含まれると推定されている[7]。

無弓類の頭蓋骨
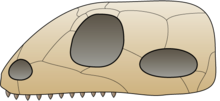
単弓類の頭蓋骨

双弓類の頭蓋骨
以下の表で記号「†」は絶滅を意味する。
爬虫類 Reptilia
- †側爬虫類 Parareptilia / 無弓類 Anapsida
- †プロコロフォン類 Procolophonia
- †パレイアサウルス科 Pareiasauridae
- †パレイアサウルス科 Pareiasauridae
- †プロコロフォン類 Procolophonia
- 真正爬虫類(階級なし) Eureptilia
双弓亜綱 Diapsida
主竜形下綱 Archosauromorpha
- †リンコサウルス類(Rhynchosauria)など
カメ目 Testudines
主竜類 Archosauria - ワニ類、鳥綱など
鱗竜類 Lepidosaurs - トカゲ、ヘビ、ムカシトカゲなど
- †側爬虫類 Parareptilia / 無弓類 Anapsida
下位分類
現生種は分類学上2つのグループ(亜目)のどちらかに所属する。これらは、頸部(首)を甲羅に納める方法によって分類される。また、その単系統性については議論があるものの[要検証]三畳紀に生息していた初期の絶滅群をプロガノケリス亜目としてまとめることもある。曲頸亜目のカメは南半球に分布し[要検証]、頸(くび)を水平に折り曲げて甲羅に納める。これに対して潜頸亜目のカメは、頸を垂直にS字形に縮めるようにして納める。また頭部では、顎を動かす筋肉である下顎内転筋が向きを変える位置も両者で異なっている。腰骨と甲羅の接続部も異なる。潜頸亜目は両者が靭帯で結合しているのに対し、曲頸亜目では双方が癒合している。
現生種は約300種とされる[8]。
- †プロガノケリス亜目
- †オドンドケリス科 Odontochelyidae :†オドントケリス Odontochelys
- †プロガノケリス科 Proganochelidae : †プロガノケリス Proganochelys
- †アウストラロケリス科 Australochelyidae: †アウストラロケリス Australochelys
- †オドンドケリス科 Odontochelyidae :†オドントケリス Odontochelys
曲頸亜目 Pleurodira
ヘビクビガメ科 Chelidae
ヨコクビガメ科 Pelomedusoidae(ヨコクビガメ科を上科とし、亜科を科として分割する説もあり[15])
潜頸亜目 Cryptodira
ウミガメ上科 Chelonioidea
ウミガメ科 Cheloniidae
オサガメ科 Dermochelyidae
- †プロトステガ科 Protostegidae :†アーケロン、等。
- カミツキガメ上科 Chelydridea
カミツキガメ科 Chelydridae

ワニガメ Macrochelys temminckii
ドロガメ上科 Kinosternoidea
メキシコカワガメ科 Dermatemydidae
ドロガメ科 Kinosternidae
リクガメ上科 Testudinoidea
ヌマガメ科 Emydidae
イシガメ科 Geoemydidae
- †メイオラニア科 Meiolaniidae :†メイオラニア、等。
オオアタマガメ科 Platysternidae
リクガメ科 Testudinidae
スッポン上科 Trionychoidea
スッポンモドキ科 Carettochelyidae
スッポン科 Trionychidae
進化
2013年理化学研究所等の国際共同研究グループによる遺伝子解読の結果、カメの祖先は約2億5000万年前の生物大量絶滅が発生した時期(P-T境界)前後にワニ、トリ、恐竜等のグループと分かれ独自の進化をした事が明らかとなった[16]。
三畳紀のカメ

プロガノケリスの全身骨格化石
(米国ニューヨークのアメリカ自然史博物館蔵)
カメ類は側頭窓を持たない。このため、鳥やワニなどよりも古く、ペルム紀に分布していた無弓類から分岐したと考えられ、祖先となった可能性のある生物として、カプトリヌス、パレイアサウルス類 (Pareiasauridae) などが挙げられていた[17][18]。
それに対し、発生学の面からは側頭窓が二次的に閉じた双弓類ではないかという説が出されていた[7]。また、脊柱の可動性など全身の骨格の形態および石灰質の卵殻を持つことなどから、カメ類は主竜類に属するのではないかという見解が示された[19][20][21]。さらに近年の分子系統学的解析の結果も、カメが双弓類それもワニや鳥など主竜類に近縁な生物であるとの見解を支持した[7][22]。この説に従えば、カメは一度は持った側頭窓を後に再び失ったと考えられる。一方、トカゲ・ヘビなどの有鱗目は、カメよりも古く分岐したとされる[13]。
カメとその他の爬虫類の中間の形態を持つ化石資料や現生生物は発見されていなかったため、カメ目の進化は極めて短期間に生じたと考えられていた[23][24]。
中国南西部の貴州省で、これまでに発見された中では最も古いカメの化石が中生代三畳紀後期、約2億2,000万年前の地層から発見された[7][25][26]。このカメは、腹側に甲羅を発達させていたが、背側は不完全であり[7]、オドントケリス・セミテスタケア(Odontochelys semitestacea、「甲羅が半分で歯のあるカメ」の意)と名付けられた。また、海生生物の化石とともに発見されたため、海生であったとされる[7]。以上の事から、初期のカメの進化の舞台が主に水中であり、下方からの捕食者の襲撃を防ぐために甲羅を発達させたとのではないかと記載者は考えている。この化石が甲羅の進化を解き明かす鍵になるかも知れないとも期待されるが、2012年現在、本種と現生のカメとの系統関係は明確になっていない。
この生物より約1,000万年後に生息していたプロガノケリスは、口蓋部に歯が残っていた点、頭部や四肢を甲内に収納できず、むしろ頭から尾の先までの全身を数多くの棘で武装していた点が現生のカメと異なるものの、現生のカメ類に近い甲羅や、嘴を持ち、側頭窓を失うという特徴をすでに具えていた[7]。プロガノケリスなど初期の属は、その他のカメ目の二亜目と姉妹グループの関係にあると考えられている[27]。
曲頸類と潜頸類の出現

約2億年前(中生代三畳紀)の世界図
ゴンドワナ大陸は南半球を中心に広がっていた。北方はローラシア大陸。
中生代の三畳紀末(約2億1,200万年前)の大量絶滅では、爬虫類や単弓類も大型動物を中心に多くの系統が絶え、当時はまだ比較的小型であった恐竜がそれ以降、急速に発展していく。カメが進化の系統上に現れるのもこの時期からである。既知で最古のカメであるオドントケリスは水生、次いで現れたプロガノケリスなどはその骨格から、陸生であったと考えられている[7]。プロガノケリス類の化石は、ドイツ、タイ、南アフリカで多数発見されており[7]、少なくともジュラ紀初期まで分布していたことが分かっている[28]。
プロガノケリス・クェンステドティ(Proganochelys quenstedti)が発見された地層からは、プロテロケルシス・ロブスタ(Proterochersis robusta)が発見されている[7]。見つかっているのは甲羅と骨盤のみであるが、腰骨が甲羅に癒合しているという特徴があった[7]。これは曲頸類と共通する特徴である[7]。そのためこれを既知で最古のヘビクビガメ類とし、曲頚類は三畳紀後期には出現していたとする説もある[7][29][要検証]。しかし、頭骨や頸椎などは発見されていないため、これが曲頸類なのか平行進化の結果なのかは判然としていない[30]。
カメはその原初から1億年の間に淡水域・陸上・海域に適応するそれぞれの系統に分岐した。現生カメ類の種の大部分を含む潜頸類が発展したのは、ジュラ紀である。最初の潜頸類とされるのは、カイェンタケリス・アプリクス(Kayentachelys aprix)で、ジュラ紀初期の地層である米国アリゾナ州のカイェンタ累層(Kayenta Formation)から出土した。この種は北アメリカ大陸最古のカメ類である[28]。
潜頸類は、ジュラ紀末には湖や川にいた曲頸類とほぼ完全に入れ替わった。また、この時期には、陸生の種も分岐したと考えられている。大規模な大陸移動の一環としてジュラ紀末にはゴンドワナ大陸が分裂したと考えられているが、このときに新しくできた海岸線によって化石種および現生種の陸棲カメの分布地域が分けられたと考える研究がある[31]
。
最初のナガクビガメ属の化石ケロディナ・アランリクシ(Chelodina alanrixi )は、オーストラリアのクイーンズランド州にある新生代第三紀始新世の地層から発見された。[32][要検証]。
ウミガメの出現
最初のウミガメは、アーケロンなどを含む潜頸亜目- ウミガメ上科- プロトステガ科(Protostegidae)である。プロトステガ科は、その形態的特徴から現生のオサガメに近い種であったと考えられている。その中でも最初の属であるサンタナケリス(Santanachelys)は、およそ1億1000万年前の白亜紀初期に現れたと考えられている。ウミガメに顕著な涙腺の肥大化という特徴は、既にこの時点で備わっていた。世界中でたくさんの化石資料が白亜紀以降の地層から見つかっている[28]。
分布

世界のカメ(カメ目)の分布 [33]
黒=カメのいる陸地 青=カメのいる水域
灰色=カメのいない陸地 白=カメのいない水域
世界の熱帯・温帯を中心に300種類ほどが分布する。一部の種は、冬眠して越冬する。また、ブランディングガメのように耐寒性を持ち、水面が凍りついた環境でも活動する種もある。海棲種は深海にまで分布を拡げる。オーストラリア大陸に現生するカメは、ウミガメとスッポンモドキ以外は曲頸亜目のカメのみである。
開発による生息地の破壊、水質汚染、食用やペット用などの乱獲などにより生息数が減少している種もいる[5]。
陸地
アフリカ大陸、オーストラリア大陸、北アメリカ大陸、南アメリカ大陸、ユーラシア大陸、インドネシア、スリランカ、セーシェル、日本、パプアニューギニア、フィリピン、マダガスカル[5]
海洋
インド洋、大西洋、太平洋、地中海[5]
日本で見られる種
外来種を含め、日本では以下の6科13種(うち2種は一部の亜種のみ)が見られる。
- ウミガメ上科
- ウミガメ科 :アカウミガメ、アオウミガメ(アオウミガメ、クロウミガメ)、タイマイ、ヒメウミガメ(上陸例なし)。
- オサガメ科 :オサガメ(上陸例なし)。
- カミツキガメ上科
- カミツキガメ科 :カミツキガメ(亜種不明)。
- スッポン上科
- スッポン科 :スッポン(= シナスッポン・ニホンスッポン)。
- リクガメ上科
- ヌマガメ科 :アカミミガメ(亜種ミシシッピアカミミガメ)。
- イシガメ科 :クサガメ、セマルハコガメ(亜種ヤエヤマセマルハコガメ)、リュウキュウヤマガメ、ニホンイシガメ、ミナミイシガメ。
発見例の少ない外来種を除く。亜種分類には諸説があるものも含む。なお、ゼニガメは本来ニホンイシガメの幼体を指す俗称であるが、クサガメの幼体もゼニガメと呼ばれる。
利用
古くから、甲羅は工芸品や占いに用いられ、肉は食用にされた。また、脂肪分は薬用、食用、美容、化粧用などの油として用いられた[9]。ウミガメ上科、アルダブラゾウガメ、ガラパゴスゾウガメ種群の脂肪、ナンベイヨコクビガメ属の卵などから採集された[9]。ガラパゴスゾウガメ種群やナンベイヨコクビガメ属の油は透明度が高く臭みも少ないことから、上質な油として欧米で取引された[9]。捕鯨船によるガラパゴスゾウガメ種群に対しての食用の乱獲は19世紀には個体数減少や、家畜の増加、灯油の普及に伴う捕鯨業者の衰退などにより減少したが、油用の乱獲は20世紀初期まで行われた[9]。
初期人類の研究では、出土するカメの骨が人類の活動が活発になるにつれて小さくなる(大型の個体群、種が食べつくされるため)ことが知られており、重要な指標となる[要出典]。
食用
日本や中国ではスッポンが古来より食用にされており、養殖もおこなわれている。イスラム圏の中央アジアや南アジアでは水棲種は不浄なものとして食用として敬遠されることもあるが、卵は食用とされることもある[34]。一方でイスラム圏でもインドネシアでは魚と同様に扱い食用とする地域もある[34]。中華人民共和国では古くから食用とされたり薬用になると信じられ、例としてミスジハコガメは癌に効く万能薬になると信じられている[35]。中華人民共和国国外に移入した中国人もカメを食用とし、アフリカ大陸、南北アメリカ大陸、東南アジア、ニューギニアなどからもカメを輸入するネットワークを構築している[34][36]。中には国際的な流通が規制されたワシントン条約附属書I掲載種が、中華人民共和国内の市場で発見された例もある[36]。大型種を乱獲しつくしたため続けて小型種を乱獲する[34]、食用や薬用で生息数が減少した種がペット用の採集や希少価値が加わることで高価になりさらに乱獲されるという連鎖現象も起こっている(Asian turtle Crisis)[36]。 香港ではミスジハコガメ(Cuora trifasciata)などの腹甲が、茯苓(ぶくりょう)などの生薬とともに煮込まれて、亀苓膏(きれいこう)、俗に「亀ゼリー」と呼ばれて、解毒・美容の効果がある食品として食べられていた。しかし、ミスジハコガメが絶滅の危機にあるため、近年は材料がクサガメなどに変えられている。弱い弾力があり、苦みと甘みがある。
オーストラリアではアボリジニが食用とし、壁画やアートのモチーフとしても用いられる[12][37]。スッポンモドキはオーストラリア国内では流通や輸出が禁止されているが、アボリジニによる自家採集は許可されている[37]。泥中で休眠するチリメンナガクビガメを人の手で泥中に埋めることで、生きた保存食としても利用された[12]。
欧米ではキスイガメが18世紀以降から大量に消費されるようになり大規模な養殖 (Turtle farming、養亀業) がおこなわれ、1880-1920年代にピークを迎えた[38]。生息数の減少と世界恐慌によりキスイガメが食用とされることは少なくなり大規模な養殖や食用の採集は行われなくなり、現在は郷土料理や家庭料理として扱われるようになった[38]。アミメガメの英名は味が鶏肉に似ていることが由来とされているが一部地域で食用とされていたとされ、食用とするのは一般的ではなく食用の養殖などが行われたこともない[39]。大航海時代にはアルダブラゾウガメやガラパゴスゾウガメ種群が食用とされ、囲心腔や膀胱に水分を溜めていることから飲水用にも利用された[9]。食物や水を与えなくても最長で2年は生存することもあり、保存食として用いられた[9]。南米では、インディオがカメを食する。
日本においても、ウミガメは貴重なタンパク源であった(特に卵は美味であるとされる)。小笠原諸島ではアオウミガメの刺身を含む各種の亀料理が発達していたが、ワシントン条約以後、捕獲が禁止されたため食用とされることはなくなった。味は鶏肉に似ている。
天然の亀は病原菌や寄生虫の宿主とされており、調理が不十分な状態での食用は危険である。生き血の飲用は特に危険であるため避けるべきである。粉末状に加工された漢方薬であっても肝炎を発症する可能性があるため、利用は避けたほうが良い。
甲羅の利用
タイマイの甲板は加工され、世界各地で工芸品の原料とされた(鼈甲)[40]。鼈甲は櫛や眼鏡のフレームなどに用いられ、日本では奈良時代から加工技術が存在する[40]。日本には1975年のワシントン条約発効後も鼈甲細工産業の保護という理由から、アメリカ合衆国による経済措置により1993年に鼈甲の輸入が禁止されるまで年あたり30トンの鼈甲が輸入されていた[40]。
ワシントン条約の附属書Iに掲載され、タイマイを含む全てのウミガメ(鼈甲製品、剥製)の国際商業取引は原則禁止されている。ただし、条約締結前あるいは付属書I掲載前に取得したものは適用除外され、また、飼育繁殖したものは附属書II掲載種と扱われるなど例外的に取引が許される場合がある[要検証]。現在は人工的に作られた鼈甲で代替されている。
古代中国の殷時代後期(3,300-3,000年前)には銅製などの刃物で亀甲や獣骨などを刻んだ亀甲獣骨文字が使用され、これは世界最古の漢字とされる[41]。殷王朝においては祭事や戦争、農耕や天気予報などに至るまで、穴をあけた亀甲や獣骨に火をあてることで生じた割れ目によって吉兆が占われる[41]亀卜が行われた。「卜」「兆」などの文字はこの際に生じた割れ目の形状に由来すると考えられている[41]。亀甲獣骨文字を刻んだ甲羅が今日まで残されている。日本ではこの亀甲による占いを「太占」と呼ぶ。
ペットの亀

「ミドリガメ」こと、ミシシッピアカミミガメ
アメリカ合衆国では、1960-1970年代にかけてアカミミガメの亜種ミシシッピアカミミガメを主とした養殖個体が大量に流通し、不衛生な環境で飼育されたことも多く動物愛護上の問題とされたりサルモネラ菌による感染症の原因(実際は食品や他の動物が原因の感染も含まれると考えられている)とみなされた[42]。そのためアメリカ食品医薬品局により1975年以降は4インチ(約10センチメートル)未満のカメの輸入や流通が規制された(教育用、実験用であれば流通可能な場合もある)[42]。
日本では「ゼニガメ」の流通名で販売されるクサガメの幼体や「ミドリガメ」の流通名で販売されるミシシッピアカミミガメなどが特に多く流通している[要検証]。それ以外にも多くの種類が流通しており、近年では日本に分布しないリクガメも飼育される。しかし、インドホシガメのように、生息地からの輸出が禁止されているにもかかわらず密輸され、流通する種もある。また、寿命が長く成長すると大型化する種類もあることから、不法な放流による環境への被害は社会問題となっている。
カメ類はペットとして広く愛好されているが、実際の飼育が必ずしも簡単とはいえない。非常に我慢強いことがペットとしての適性にかなっていると誤解されている面もある。さらに適切な飼育環境下では長寿なこともあり、長期飼育を視野に入れる必要がある[4]。
動物行動学者コンラート・ローレンツはヨーロッパでペットとしてよく飼育されたギリシャリクガメについて、たいていの場合、死ぬのに時間がかかっているだけと酷評し、良好な条件で飼育することの困難さを指摘している[43]。
文物に登場する亀
神話・伝説の亀

湖北省博物館所蔵の玄武像
15世紀初期の作
カメは地域や民族によっては信仰の対象とされる。中国文明影響下の東アジア文化圏では、霊獣四神の一つとして玄武があり、北方を守護するとされた。脚の長い亀に蛇が巻き付いた形で描かれることが多いが、尾が蛇となっている場合もある。先述のように古代中国ではカメの甲羅を用いて神託の儀式を行っていた。『史記』中にも神聖な亀について一書がある。
バラモン教による古代インドの人々の宇宙観では、複数で大地からなる我々の世界は3頭の巨象に支えられ、その巨象たちは1頭の大亀に支えられている。そしてさらには、その大亀は1匹の未曾有の体躯を持つ大蛇の上に乗っている [要出典]。ヒンドゥー教の時代、ヴィシュヌ信仰が盛んになると先の大亀はその化身であるところの大亀クールマであるとされた。世界(その一つ)であるマンダラ山が大蛇ヴァースキによって海底に沈められようとしたときに、大神ヴィシュヌが大亀の姿となって山を背に乗せ、それによって世界は保たれることになったという。また、クールマより古い亀の王アクーパーラも同様、マンダラ山を支える偉大な大亀で、本質的に同一である。
ギリシャ神話では変身譚の一つとしてケローネーの物語がある。ケローネーはニュンペーの一人で、ゼウスとヘーラーとの結婚式を馬鹿にして出席しなかったことから神々の怒りを買い、ヘルメースによって亀の姿に変えられる。「ケローネー」(Chelone)はギリシャ語で「亀」を意味する言葉である。

怪物タラスクの像(南フランスのタラスコン。これは観光用に作られたものであるが、姿形がよく分かる)
フランス南部の町タラスコンには、亀に似た怪物タラスクの伝説が伝わっている。この怪物は硬い甲羅に鋭い背鰭とヤマネコの上半身、6本の肢とドラゴンの体を具えている。
日本では亀の登場する伝説として、『浦島太郎』の物語がある。海辺で子供にいじめられていた海亀を助けた太郎は、この亀によって海底にある竜宮城へ連れて行かれそこで乙姫からもてなしを受ける。しばらくたって浦島が地上へ帰ろうとすると、乙姫は「決して開けてはならない」と注意しつつ土産として玉手箱を渡す。浦島が地上に戻ると、彼のことを知っている人が誰も見つからない。そこで浦島が玉手箱を開けると、そこから出てきた煙を浴びて、浦島は老人になってしまう。一般的にはこのような筋として知られているが、典拠によって様々なバリエーションがある(浦島太郎を参照)。
別伝であるが、乙姫が病気になり、竜宮の王が薬となる猿の肝を手に入れる必要が生じた、という話がある。その際、猿を呼び寄せるために、陸にも出られるからとカメが使いをする。カメは猿をだまして海に連れて行くが、途中で理由を漏らし、猿が「肝は枝に干してある」というのを信じて逃がしてしまう。カメは罰として甲羅を割られたと、これは甲羅の形の由来譚である。
吉兆・縁起物としての亀

『名所江戸百景 深川 萬年橋』 歌川広重 画
日本では「鶴は千年 亀は万年」と言われ、鶴とともに亀は長寿の象徴、夫婦円満の象徴とされる。夫婦円満の象徴とされるのは、先述の『浦島太郎』伝説の原典の一つにおいて、浦島太郎が老人になったのちも乙姫(亀姫)が太郎を慕い続けて添い遂げ、やがて太郎は鶴に、乙姫は亀に化身したというところから来ている[要出典]。「亀姫」など名前としても使われる。
浮世絵師・歌川広重の『名所江戸百景 深川 萬年橋』(右方の画像を参照)には、手桶の取っ手に吊るされたニホンイシガメであろう1匹の亀が描かれているが、これは画題「萬年橋」の「萬年(万年)」を「鶴は千年 亀は萬年」にかけたもの。近景の亀も、手桶の取っ手と窓枠が形作る額のような四角画面に納まった遠景の富士の山も、そして2艘の帆掛け舟も、みな縁起物である。

『木曽街道六十九次之内 福島 浦島太郎』 歌川国芳(画号:一勇斎国芳)画
緑藻類の付着したカメのことのことを蓑亀とよぶ。背中に蓑を羽織ったように見えることに由来し、日本では他に「緑毛亀」「緑藻亀」などと呼ばれる。中国や日本では長寿を象徴する縁起のよいものとして珍重され、古くからさまざまな文学作品や芸術作品に記述が見られる。
「動きの遅いもの」としての亀
地中海文明および西欧文明においては、『イソップ寓話』中の一話「ウサギとカメ」が有名である。ここでの亀は鈍重で一見無能と思われながら、歩みは遅くとも着実に真っ直ぐ進む、いわゆる「勤勉さ」の象徴として描かれている。
古代ギリシアの無限に関する哲学的パラドックスとして有名な「アキレスと亀のパラドックス」では、俊足で知られる英雄アキレスに対して、歩みの遅いものの象徴として亀が用いられている。「先行する亀にアキレスが永遠に追いつけない」という常識的に受け入れがたい結論を導くパラドックスであり、アキレスに比べ亀が遅いものであるとの共通認識が無ければそもそもパラドックスとして成り立たない。
日本海軍においては潜水艦を水上艦船と比較して、「どん亀」と揶揄する表現があった。
変速機構においてHi-Loの意味でウサギとカメの絵が描かれる例は多く存在する。例えば建設機械の変速機構操作にはJIS規格としてウサギとカメの絵が描かれている[44]ほか、ミシンの速度調節としてもウサギとカメが用いられている。
侮蔑表現としての亀
現代中国においては亀は侮蔑表現の一つとされる。中国語の最大の侮蔑語に「王八蛋(ワンパータン)」というものがあるが王八が亀を意味し中国人が守るべき八徳(仁・義・礼・智・信・忠・孝・悌)を忘れるという意味の「忘八(ワンパー)」と発音が似ているため、「王八蛋」で(八徳を忘れてしまうような)愚か者、恥知らずの意味として用いられる。
近現代の創作の題材としての亀
- 『ガメラ』シリーズ :1965年初出の大映(現・角川映画)による怪獣映画。『ゴジラ』のヒットに影響されて製作されたものが最初であるが、怪獣ガメラは、当時の子供がペットとして夢中になっていたミドリガメをモチーフとし、空飛ぶ円盤(モチーフ)との組み合わせ、並びに、(いかにも亀らしく)子供の味方との設定も功を奏して、以後長くシリーズを重ねることになる一大キャラクターとして登場した。視聴者の年齢層が上がった(初期の視聴者層が大人になった)平成以降のシリーズでは、ミドリガメに変えて“強面(こわもて)の”ワニガメが造形モチーフとされている。なお、設定上、昭和ガメラはアーケロンを祖先に持つとのことである。また、新たに発見された古代カメの一種が飛行機の後退翼に酷似した突起物を甲羅の側辺に具えていることから空飛ぶカメであるガメラを連想し、「シネミス・ガメラ(Sinemys gamera、『中国の亀、ガメラ』の意)」という学名が与えられている。
- 漫画 『ティーンエイジ・ミュータント・ニンジャ・タートルズ(忍者タートルズ)』 :1984年初出の米国作品。ミュータントで擬人化された4匹の架空の亀の忍者のグループが主人公。のろまな冷血の爬虫類と素早く技巧的な日本の武術とのミスマッチによる非常に滑稽な印象がデザインの元となった。
- ゲーム 『カメ一族』 : 任天堂『マリオシリーズ』に登場するノコノコをはじめとする架空のキャラクター群。初出である1985年のスーパーマリオブラザーズ以降、ほぼ全作に登場している。「下から叩いてもやられなくて、やがて復活してくる」とのコンセプトからカメが採用された[45]。ボスキャラクターであるクッパは「テレビゲーム史上最も偉大な敵役」としてギネスに登録されている[46]。マリオカートなど、甲羅が単独で出現する場合もある。
呼称
学名(ラテン語)
ラテン語では「カメ」を testūdō (テストゥードー)と言う[47]。これは陶製の容器testūや、煉瓦や土製の容器testaに由来する[47]。testūdōの複数形 testūdinēs (テストゥーディネース)が生物群としての「カメ類」の呼称、そして、和名の「カメ目」にあたる学名 Testudines ともなっている。
これとは別に、古典ギリシア語χελώνηに由来するchelysもカメを意味する[48]。この語は(亀の甲で作った)竪琴も指すためChelysはこと座を意味する[48]。
漢字文化圏

漢字の原形を色濃くとどめる中国の字体(「新字形」の繁体字)。なお、画像は亀苓膏(亀ゼリー)などを食べさせる中国の店(武漢市。「#食用の亀」)。
古代中国文明影響下の東アジアにおいて、亀の甲羅は、儀式に則り火に炙ったときの割れ方で神意を占うものであった[49]。
漢字の「亀」は古代中国の亀甲獣骨文字(甲骨文字)に起源し、元は亀の体形を真上から描いた象形文字であった[50]。現在の文字と直結する造形は秦代の小篆に見ることができるが、ここでの亀は側面から描かれ、角度を縦に変えられている。詳しくはウィクショナリーの「龜」、および、その一節「字源」を参照。中国の繁体字(新字形)は現代のものでも、なお原形に近い(「他の言語」の「中文」、および、右の画像を参照)。頭と胴体、四肢と尾が揃っており、背中に甲羅を背負っている。地域などによって字形にかなり差異がある(繁体字#対照例示参照)。
現代中国語では陸棲、淡水棲のものを「烏龜」(wūguī、ただしクサガメの別名としても使われる)[51]、海棲のものを「海龜」(hǎiguī)という。陸生のもののうちスッポンについては「鱉(鼈)」(biē)または「甲魚」(jiǎyú)[51]などという。
英語
英語圏ではウミガメ上科はturtle、リクガメ科はtortoiseと呼称され、淡水棲種を指す1語はなくfreshwater turtleと呼称される[9][39]。一方でカメ目全体の総称としてはアメリカ合衆国ではturtles、イギリスではturtle & tortoiseを用いることが多い[39]。
アメリカ合衆国では陸棲種をterrestrial turtleと呼称することもあり、特にリクガメ科を除く陸棲種を指して用いられることもある[39]。キスイガメの種小名terapinおよび英名はアルゴンキン語族の「食用ガメ」を指す語句に由来する[39]。アメリカ合衆国ではキスイガメのみをterrapinと呼称するのが一般的で、クーターガメ属をcooter、アカミミガメ属をsliderなどのように特定の分類群を指す単語を用いることが多い[7]。
イギリスではturtleはウミガメ上科のみを指すという動きが強く、統一がとれていないものの淡水棲種をterrapinと呼称することもある[39]。ただし、例えば "Fly River turtle" のように、北米やオーストラリアで広く知られている名称があれば、例外としてそれを用いる。
オーストラリアでは水棲種はturtle、リクガメ科はtortoiseと呼称される[39]。オーストラリアにはリクガメ科などの陸棲種がいないため淡水性種もtortoiseと呼称する動きも過去にあったが、一般的ではない[39]。
べっこう(鼈甲)はウミガメ(sea turtle)の一種タイマイから作られるのに、tortoiseshell(直訳「リクガメの甲羅」)と呼ばれている。そのためタイマイ(hawksbill turtle)はtortoiseshell turtleとも呼ばれることがある。このように英語におけるtortoiseとturtleの使い方は混乱している。
tortoiseはラテン語torquēre(直説法現在一人称単数形torqueō、「ねじる」「曲げる」[52])に由来する語根「tort-」から、足のねじくれた動物という意味である[53]。よってtorsion(「ねじれ」)やtorture、torment(ともに「捻じ曲げる」、「拷問にかける」)と同系[53]。turtleも語源はtortoiseと同じである。
混乱を避けるためにカメ目すべてを表せる単語として chelonian を用いることが、英語圏の獣医師や科学者、自然保護論者を中心に支持されている[要出典]。chelonian は、古代ギリシア語で "tortoise" を表す χελώνη (khelōnē、ケローネー。現代ギリシャ語の χελώνα)から来ている。
ドイツ語
ドイツ語ではカメのことをSchildkröteと呼ぶ。 ドイツ語では複数の単語をつないで一単語を形成することが多いが、Schildは英語のshieldに相当する単語で、甲羅を意味する。一方、Krötenはヒキガエルの意味がある。
タイ語
タイ語ではเต่า(tao、タウ)といい、タイ文字の「ต」の名称「ต เต่า」(to tao、トータウ、意味は「亀のトー」)となっている。
亀に関する言葉
- 「亀鳴く」
- 春の季語として「亀鳴く」という言葉がある。『夫木集』に収められた藤原為家の歌、「川越のをちの田中の夕闇に何ぞと聞けば亀のなくなり」に由来するとされ、古くから季語として定着している[54]。なお「亀の子」は夏の季語である。
- 亀甲
- 亀の甲羅のことであるが、漢字文化圏での言葉としては、亀の甲羅の模様に似た六角形のことを喩えて言う。
- 亀の甲より年の劫
- 「劫(こう)」は極めて長い時間。年長者の経験や知恵は尊ぶべきであるということの喩えで、「甲(こう)」と「劫(こう)」を音でかけたもの(日本)。「蟹(かに)の甲より年の劫」も同義。また、意味からは「医者と坊主は老人がよい」や「松傘より年嵩(としかさ)」も同じ。
- 亀甲縛り
亀甲縛りは戦国時代から江戸時代にかけて発達を見せた日本式捕縄術中の一手(「緊縛#全身の縛り方」参照)。縛り上げた結果、縄目が亀の甲羅の模様に似た六角形の組み合わせ図形を描き出すことからこの名がある。現在ではSMにおける緊縛でよく知られる。- 亀船(亀甲船)
亀船(亀甲船)は、文禄・慶長の役で活躍したと伝えられる李氏朝鮮の軍艦。守りの堅い構造上の特徴、意匠的主旨ともに亀と亀甲に通じている。
- 亀石
亀石は、日本における巨石文化の古代遺跡の一つであり、亀の形に似ていることから古来そのように呼ばれていたものと考えられる。ただし、この遺跡が何を目的に造られたものであるのかはまったくの謎である。- 月と鼈
- 月と鼈(すっぽん)はともに形は丸くて一見似ているが、その実まったく違うものであることから、比較にならないほどかけ離れていることの喩え(日本)。多くの場合、月を優れたもの、鼈を劣っているものと捉えて比較の意味合いをもって使われる。
- 鶴亀算
鶴亀算は中国の数学書を参考に江戸時代に考案され、広く用いられていた日本の応用算術。2本の足を持つ鶴と、4本の足を持つ亀を想起しながら文章題を解くものであり、吉兆の象徴ゆえに鶴と亀が選ばれている。

亀3匹。
- 親亀の背中に子亀を乗せて
- 「親亀の背中に子亀を乗せて そのまた背中に孫亀乗せて そのまた背中に曾孫(ひ-まご)を乗せて 親亀こけたら皆こけた」 日露戦争の戦勝気分に沸く明治末期の日本で大流行し、今でも歌われる、『ラッパ節(音声資料:[55])』第11番の歌詞。
- 格闘技の「亀」
- 格闘技においては、うつ伏せになり脇を閉め顎を引いた防御姿勢のことを亀という。特に柔道で多用される。詳細は亀 (ガードポジション)を参照。
- 亀の子たわし
- 「亀の子たわし(亀の子束子)」は、椰子(やし)の繊維を加工して作られた、日本に独特の洗浄用具であり、その形状が亀の子供によく似ていることから名づけられたものである。1907年(明治40年)考案。
- タートルネック
- 「タートルネック」は、亀の頸もとの様子に似て、首の密着する丸くて高い襟(えり)のことである。これはアメリカ英語であるが、日本では1960年代後期から70年代前期にかけて流行し、言葉としても普及した。また、当時、日本語で「徳利首(とっくり-くび)」とも呼んでいた。
- 亀頭
- 「亀頭(きとう。陰茎亀頭)」は、男性器の先端部位の形状が亀の頭部および頸部の表皮の様子に酷似していることから、そのように名付けられた日本語である。
カメの名を持つ生物
日本語、その他言語のカメ(亀、turtleなど)の名を冠する、または含む生物を挙げた。普通、カメの姿またはその一部分に似ていることに由来する。スッポン(鼈甲など)を由来とするものは省いた。
和名

カメガエル。カメに似ている。
原則五十音順となっている。
オオカメノキ - 学名Viburnum furcatum。スイカズラ科ガマズミ属の落葉樹。葉の形を亀の甲羅に見立てて「大亀の木」と呼ばれる。ただし異説もある[56]。
カメガイ - 軟体動物門腹足綱後鰓類有殻翼足目のカメガイ科に属する巻貝の一種または総称。薄い殻が亀の甲のような形をしているため[57]。
カメガエル - 学名Myobatrachus gouldii。両生綱無尾目カエル亜目カメガエル科に属するカエルの一種。カメの姿に似ていることから。英名もカメに由来するTurtle Frog。- 「カメノコ」を冠するもの - 「亀の子」、または「亀の甲」に由来する。
カメノコキクメイシ - 学名Favites abdita。刺胞動物門花虫綱イシサンゴ目キクメイシ科に属するサンゴの一種。亀の甲に似た模様から。別属のものも「カメノコキクメイシ」の名を含むものがある。
カメノコデオキノコムシ - 学名Cyparium mikado。鞘翅目デオキノコムシ科の昆虫。
カメノコテントウ - 学名Aiolocaria hexaspilota。鞘翅目多食亜目テントウムシ科に属するテントウムシの一種。翅は黒地に橙色の模様があり、これが亀の甲の模様に似ていることから。
カメノコハムシの仲間 - 鞘翅目多食亜目ハムシ科カメノコハムシ亜科に属する昆虫。危険を感じると脚と頭を体の中に隠し、それがカメのようであることから。代表種はカメノコハムシ Cassida nebulosaなど。英名もカメに由来するTortoise Beetle。
カメノコフシエラガイ - 学名Pleurobranchus peroni。軟体動物門腹足綱側鰓目カメノコフシエラガイ科ゼニガタフシエラガイ属のウミウシの一種。
カメノコロウムシ - 学名Ceroplastes japonicus。半翅目カタカイガラムシ科ロウムシ属に属するカイガラムシの一種。蠟物質に覆われている。
カメノコウムシ - 学名Gallerucida bifasciata。鞘翅目多食亜目ハムシ科のイタドリハムシの別名。亀の甲から。
カメノコウワムシ - 学名Keratella cochlearis。輪形動物門遊泳目ツボワムシ科。殻に「亀の甲」模様があることから[58]。
カメノテ - 学名Capitulum mitella。節足動物門顎脚綱蔓脚下綱無柄目ミョウガガイ科カメノテ属の甲殻類。見た目の形状がカメの手に似ていることから。
カメバス - 学名Nymphaea tetragona。スイレン科スイレン属の水生植物ヒツジグサの別名。意味は「亀蓮」。
カメバヒキオコシ - 学名Isodon umbrosus var. leucanthus f. kameba。シソ科ヤマハッカ属に属する植物、イヌヤマハッカの亜種。葉は先端が3裂し、その中央をカメの尾と見立てて名づけられた[59]
カメフジツボ - 学名Chelonibia testudinaria。節足動物門顎脚綱蔓脚下綱有柄目フジツボ亜目オニフジツボ超科に属する甲殻類。見た目ではなく、ウミガメに付着することから。
カメホウズキチョウチン - 学名Terebratalia coreanica。腕足動物門嘴殻亜門嘴殻綱穿殻目に属するチョウチンガイの一種(貝ではない)。
カメムシの仲間 - 半翅目カメムシ亜目に属する昆虫の総称。形状が亀の甲に似ていることから[60]。ただし瓶に似ているからという説もある[61]。- 「キッコウ」(亀甲)を冠するもの - 亀甲の形、もしくは紋様に見立てられるもの。
キッコウカンザン - 学名Pleioblastus hindsii f. kiko。イネ科タケ亜科メダケ属カンザンチクの一品種。桿の基部で節間が亀甲状になるため。
キッコウグサ - 学名Dictyosphaeria cavernosa。アオサ藻綱シオグサ目バロニア科に属する藻類。
キッコウゴケ - 学名Diploschistes actinostomus。子嚢菌門チャシブゴケ綱オストロパ目キッコウゴケ科キッコウゴケ属に属する地衣類。同属にウスグロキッコウゴケ Diploschistes aeneusなどがある。
キッコウソウ - 学名Pyrola japonica。ツツジ科イチヤクソウ属の多年草、イチヤクソウの別名。
キッコウダカラ - 学名Mauritia maculifera。タカラガイ科タカラガイ属の巻貝の一種。
キッコウダコ - 学名Alocasia cuprea。サトイモ科クワズイモ属に属する植物。
キッコウツゲ - 学名Ilex crenata var.nummularia。モチノキ科モチノキ属の常緑低木イヌツゲの変種。
キッコウハグマ - 学名Ainsliaea apiculata。キク科モミジハグマ属の多年草。葉が五角形で、これを亀甲に見立てた[62]。
キッコウヒイラギ - 学名Osmanthus heterophyllus f. subangustatus。モクセイ科モクセイ属ヒイラギの園芸品種。
キッコウモンケシカミキリ - 学名Exocentrus tetsudineus。鞘翅目カミキリムシ科フトカミキリ亜科に属するカミキリムシの一種。上翅に白色毛による亀甲紋がある。種小名tetsudineusもカメに由来する。
キッコウリュウ - 学名Dioscorea elephantipes、英名elephant's foot。ヤマノイモ科ヤマノイモ属に属する植物。
英名
和名で挙げたものを除く。
Turtle Dove - キジバトの仲間。ハト目ハト科に属する。単にTurtle doveといった場合特にコキジバト Streptopelia turturを指す。翼の模様が亀の甲に似ていることから。
Turtle Grass - 学名Thalassia testudinum。トチカガミ科リュウキュウスガモ属の海草。
Turtlehead - オオバコ科(旧ゴマノハグサ科)ジャコウソウモドキ属の多年草、ジャコウソウモドキの仲間。花の形がカメの頭に似ていることに由来する。属名Cheloneも古代ギリシア語でカメを表す χελώνηに由来する。
画像

ギリシャリクガメ
Testudo graeca
(リクガメ科)

スッポンモドキ
Carettochelys insculpta
(スッポンモドキ科)

アオウミガメ
Chelonia mydas
(ウミガメ科)

マタマタ
Chelus fimbriatus
(ヘビクビガメ科)

カミツキガメ
Chelydrta serpentina
(カミツキガメ科)

オサガメ
Dermochelys coriacea(オサガメ科)

ヨーロッパヌマガメ
Emys orbicularis
(ヌマガメ科)

スペングラーヤマガメ
Geoemyda spengleri
(イシガメ科)

ヌマヨコクビガメ
Pelomedusa subrufa
(ヨコクビガメ科)

オオアタマガメ
Platysternon megalocephalum
(オオアタマガメ科)

オドントケリス
Odontochelys semitestacea










脚注
^ 近年の分類による。詳しくは「#上位分類」を参照。
- ^ ab“甲羅のないカメの化石を発見 進化解明に貢献”. BBC. (2018年8月24日). https://www.bbc.com/japanese/45280112 2018年9月1日閲覧。
^ 松井編(2006),p.134
- ^ abcde安川雄一郎 「ビギナーにおすすめのカメ12種〜初心者向けとして飼育者に薦めるカメ類〜」『エクストラ・クリーパー』No.1、誠文堂新光社、2006年、113-122頁。
- ^ abcdefghijklmnopqrstuvwxyzaaabacad深田祝監修 T.R.ハリディ、K.アドラー編 『動物大百科12 両生・爬虫類』、平凡社、1986年、85-99頁。
^ 安川雄一郎 「ペットとしてのリクガメの飼育と分類」『エクストラ・クリーパー』No.3、誠文堂新光社、2008年、56-57頁。
- ^ abcdefghijklmnopqrstu安川雄一郎 「オオアタマガメの分類と自然史」『クリーパー』第45号、クリーパー社、2008年、24-28頁。
- ^ abcdefghijklmnopqrstuvwxyz安川雄一郎 「水棲ガメの世界」『ハ・ペト・ロジー』Vol.3、誠文堂新光社、2005年、14-48頁。
- ^ abcdefghijklmn安川雄一郎 「ゾウガメと呼ばれるリクガメ類の分類と自然史(後編)」『クリーパー』第33号、クリーパー社、2006年、16-29、32頁。
^ 胚発生過程と化石記録から解き明かされたカメの甲羅の初期進化 独立行政法人理化学研究所プレスリリース 2013年7月9日
^ Anatomy and Diseases of the Shells of Turtles and Tortoises
- ^ abc柴田弘紀 「ナガクビガメ総覧」『エクストラ・クリーパー』No.1、誠文堂新光社、2006年、42-45頁。
- ^ ab[リンク切れ]「カメの系統的位置と爬虫類の進化-宮田隆の進化の話」 『JT生命誌研究館』, 2008年9月26日 (金) 19:43 UTC、URL: http://www.brh.co.jp/katari/shinka/shinka04.html
^ 金子隆一 著 『哺乳類型爬虫類 : ヒトの知られざる祖先』 P32
^ 安川雄一郎 「アフリカヨコクビガメ亜科の分類と自然史 その1」『クリーパー』第34号、クリーパー社、2006年、25-26頁。
^ ゲノム解読から明らかになったカメの進化 -カメはトカゲに近い動物ではなく、ワニ・トリ・恐竜の親戚だった- 理化学研究所 報道発表資料 2013年4月29日
^ LEE, M. S. Y. 1995. Historical burden in systematics and the interrelationships of ‘parareptiles’. Biological Reviews, 70, 459–547.
^ LEE, M. S. Y. 1997. Pareiasaur phylogeny and the origin of turtles. Zoological Journal of the Linnean Society, 120, 197–280.
^ deBRAGA, M. and RIEPPEL, O. (1997), Reptile phylogeny and the interrelationships of turtles. Zoological Journal of the Linnean Society, 120: 281–354.
^ RIEPPEL, O. 1997. Reptile phylogeny and the interrelationships of turtles. Zoological Journal of the
Linnean Society, 120, 281–354. DE BROIN, F. 1984. Proganochelys ruchae
^ HEDGES, S. B. and POLING, L. 1999 A molecular phylogeny of reptiles. Science, 283, 898–901.
^ 平山廉 著 『カメのきた道』61 - 64ページ
^ Nagashima, Hiroshi; Shigehiro Kuraku, Katsuhisa Uchida, Yoshie Kawashima Ohya, Yuichi Narita and Shigeru Kuratani (May 2007). “On the carapacial ridge in turtle embryos: its developmental origin, function and the chelonian body plan”. Development (134): 2219-2226. doi:10.1242. http://dev.biologists.org/cgi/content/abstract/134/12/2219.
^
「カメはどのようにして甲羅を獲得したのか」 『理研CDB-科学ニュース』 2008年9月26日 (金) 19:43 UTC、URL: http://www.cdb.riken.go.jp/jp/04_news/articles/pdf/070611_carapace_approved.pdf
^ “甲羅進化の謎に迫る最古のカメを発見”. ナショナルジオグラフィック. 2008年11月26日閲覧。
^ “最古のカメ類化石が発見 進化の謎を解く大きな手がかり”. 人民網. 2008年11月28日閲覧。
^ Gaffney and MEEKER, L. J. 1983. Skull morphology of the oldest turtles: a preliminary description of Proganochelys quenstedti. Journal of Vertebrate Paleontology, 3, 25–28.
- ^ abcP.M. Datta; P. Manna , S.C. Ghosh & D. P. Das (Nov 2003). “The First Jurassic turtle from India”. Palaeontology 43 (1): 99 - 109. doi:10.1111/1475-4983.00120. http://www3.interscience.wiley.com/cgi-bin/fulltext/119037378/PDFSTART.
^ FRAAS, E. 1913. Proterochersis, eine Pleurodire Schildkrote aus dem Keuper. Jahreschefte des Vereins fur Vaterladische Naturkunde in Wu¨rttemberg, 79, 13–30.
^ 平山廉 著 『カメのきた道 : 甲羅に秘められた2億年の生命進化』79 - 80頁
^ Hirayama, Ren; Donald B. Brinkman, and Igor G. Danilov (2000). “DISTRIBUTION AND BIOGEOGRAPHY OF NON-MARINE CRETACEOUS TURTLES”. Russian Journal of Herpetology 7 (3): 181 - 198.
^ Lapparent de Broin, F. de, & Molnar, R. E., 2001: Eocene chelid turtles from Redbank Plains, Southeast Queensland, Australia. –Geodiversitas: Vol. 23, #1, pp. 41-79
^ Cogger, H.G & Zweifel, R.G. (1998). "Reptiles & Amphibians". ISBN 0121785602
- ^ abcd安川雄一郎 「バタグールガメ属の分類と生活史1」『クリーパー』第28号、クリーパー社、2005年、105、109-110頁。
^ 安川雄一郎 「アジアハコガメ属の分類と生態・生活史1」『クリーパー』第4号、クリーパー社、2000年、11-14頁。
- ^ abc安川雄一郎 「セダカガメ類(Pangshura、Kachuga)の分類と生活史」『クリーパー』第27号、クリーパー社、2005年、17-18頁。
- ^ ab安川雄一郎 「スッポンモドキの分類と自然史」『クリーパー』第24号、クリーパー社、2004年、8-23頁。
- ^ ab安川雄一郎 「キスイガメの分類と生活史1」『クリーパー』第47号、クリーパー社、2009年、40-41頁。
- ^ abcdefgh安川雄一郎 「アミメガメ属の分類と自然史」『クリーパー』第46号、クリーパー社、2009年、82-87、90-105頁。
- ^ abc小原秀雄、浦本昌紀、太田英利、松井正文編著 『動物世界遺産 レッド・データ・アニマルズ5 東南アジアの島々』、講談社、2000年、106-107、204頁。
- ^ abc伊藤通弘、「紙の発生から普及まで (5)」『紙パ技協誌』 1996年 50巻 1号 p.200, doi:10.2524/jtappij.50.200, 紙パルプ技術協会、
- ^ ab安川雄一郎 「アカミミガメ属(スライダーガメ属)の分類と自然史1」『クリーパー』No.36、クリーパー社、2007年、46-48頁。
^ コンラート・ローレンツ 『ソロモンの指環』 日高敏隆訳、ハヤカワ文庫
^ 2014年5月13日放送有吉弘行のダレトク!?
^ “社長が訊く『New スーパーマリオブラザーズ Wii』”. 任天堂. 2014年5月24日閲覧。
^ GuinnessWorldRecords.com (2013年1月23日). “Bowser crowned ‘greatest videogame villain of all time’ in poll for Guinness World Records 2013 Gamer's Edition”. 2014年5月24日閲覧。
- ^ ab水谷智洋『改訂版 羅和辞典』研究社、初版1952年9月、ISBN9784767490250、660頁。
- ^ ab水谷智洋『改訂版 羅和辞典』研究社、初版1952年9月、ISBN9784767490250、115頁。
^ たとえば韓非子・初見秦篇に「襄主鑽龜筮占兆、以視利害、何國可降」[1]、淮南子・時則訓には「是月、命太祝、禱祀神位、占龜策、審卦兆、以察吉凶」[2]などとある。
^ 説文解字「象足甲尾之形」[3](足・甲・尾の形に象る)。
- ^ ab欧米・アジア語学センター編『現代中国語会話辞典』、日東書院、2007年、149頁。
^ 水谷智洋『改訂版 羅和辞典』研究社、初版1952年9月、ISBN9784767490250、671頁。
- ^ ab田代正雄『語源中心英単語辞典』南雲堂、2005年3月9日改装1刷、ISBN9784523310426、294頁。
^ 角川春樹編 『合本現代俳句歳時記』 角川春樹事務所、1998年、221頁。
^ ラッパ節(原曲) :音量注意。
^ “オオカメノキ 【六甲山系植生電子図鑑】”. 国土交通省 近畿地方整備局 六甲砂防事務所. 2014年7月10日閲覧。
^ “『世界大百科事典 第2版』カメガイ”. kotobank. 2014年7月10日閲覧。
^ 一瀬諭・若林徹哉『普及版 やさしい日本の淡水プランクトン図解ハンドブック 改訂版』合同出版、2008年10月10日改訂版発行、104頁
^ 杉村昇『名前といわれ 野の草花図鑑2』偕成社、1985年4月発行、92頁
^ 加納喜光『動植物の漢字がわかる本』山海堂、2007年1月10日発行、99頁。
^ 大谷剛、栗林慧『名前といわれ 昆虫図鑑』偕成社、1987年7月発行、ISBN9784035290506、38頁、104頁
^ 佐竹義輔・大井次三郎・北村四郎他編『日本の野生植物 草本III合弁花類』、1981年、平凡社。
参考文献
疋田努 『爬虫類の進化』東京大学出版会、2002年、ISBN 4-13-060179-2。- 平山廉 著、小田隆 復元画 『図解雑学:恐竜の謎』 ナツメ社、ISBN 4-8163-3306-1。
- 平山廉 『カメのきた道 : 甲羅に秘められた2億年の生命進化』 NHKブックス、ISBN 978-4-14-091095-5。
- 岩槻邦男・馬渡峻輔監修;松井正文編集、『脊椎動物の多様性と系統』,(2006),バイオディバーシティ・シリーズ7(裳華房)
関連項目
- 爬虫類
外部リンク
疋田努、千石正一 「日本産爬虫両生類の和名の変遷と現状」 『爬虫両棲類学会報』 Vol.2000 (2000) No.1 P.20-33- 太田英利、高橋亮雄 「カメの分類: 特に邦産種の学名の変更を中心に」 『爬虫両棲類学会報』 Vol.2006 (2006) No.2 P.131-139
- 太田英利 「スタイネガー (1907) に掲載された日本とその周辺地域のカメ類」 『爬虫両棲類学会報』 Vol.2007 (2007) No.2 P.203-211


